

The basic function of the gallbladder in humans is one of protection. The accumulation of the primary bile acids (cholic acid and chenodeoxycholic acid) in the gallbladder reduces the formation of the secondary bile acids (deoxycholic acid and lithocholic acid), thus diminishing their concentration in the so-called gallbladder-independent enterohepatic circulation and protecting the liver, the stomach mucosa, the gallbladder, and the colon from their toxic hydrophobic effects. The presence or absence of the gallbladder in mammals is a determining factor in the synthesis of hydrophobic or hydrophilic bile acids. Because the gallbladder contracts 5-20 min after food is in the stomach and the “gastric chyme” moves from the stomach to the duodenum 1-3 h later, the function of the gallbladder bile in digestion may be insignificant. The aim of this article was to provide a detailed review of the role of the gallbladder and the mechanisms related to bile formation in humans.
La función básica de la vesícula biliar en el ser humano es de protección. Mediante la acumulación de los ácidos biliares primarios (ácidos cólico y quenodesoxicólico) en la vesícula biliar, se disminuye la formación secundaria de ácidos biliares hidrófobos tóxicos (ácidos desoxicólico y litocólico), lo que reduce su concentración en la vesícula de forma independiente de la circulación enterohepática y protege al hígado, la mucosa del estómago, la vesícula biliar, y el colon de sus efectos. En los mamíferos la presencia o ausencia de la vesícula biliar es determinante para la síntesis de ácidos biliares hidrófobos o hidrófilos, respectivamente. Debido a que la vesícula biliar se contrae 5-20min después de que los alimentos están disponibles en el estómago, y «el quimo gástrico» se mueve desde el estómago hasta el duodeno solamente 1-3h más tarde, la función de la vesícula biliar en la digestión es realmente mínima. El objetivo de esta revisión es hacer una revisión detallada del papel de la vesícula biliar y los mecanismos relacionados con la formación de bilis en el humano.
The prevalent point of view is that the gallbladder is not essential for life.1 The gallbladder has the functions of absorption, concentration, secretion, and evacuation.2,3 The absorption and concentration functions are interdependent. The absorption function of the gallbladder includes the absorption of water, Na+, cholesterol, phospholipids, hydrophilic proteins, etc.4–14 Since the absorption of the bile acids by the gallbladder mucosa is 2–6% of the total concentration in the gallbladder bile, the concentration function of the gallbladder consists of accumulating the bile acids from the hepatic bile in the gallbladder.10–12,15,16 The secretion function of the gallbladder includes the secretion of glycoprotein mucin by the gallbladder mucosa, H+ ions, Cl−, and probably immunoglobulins and Ca2+.5,17–23
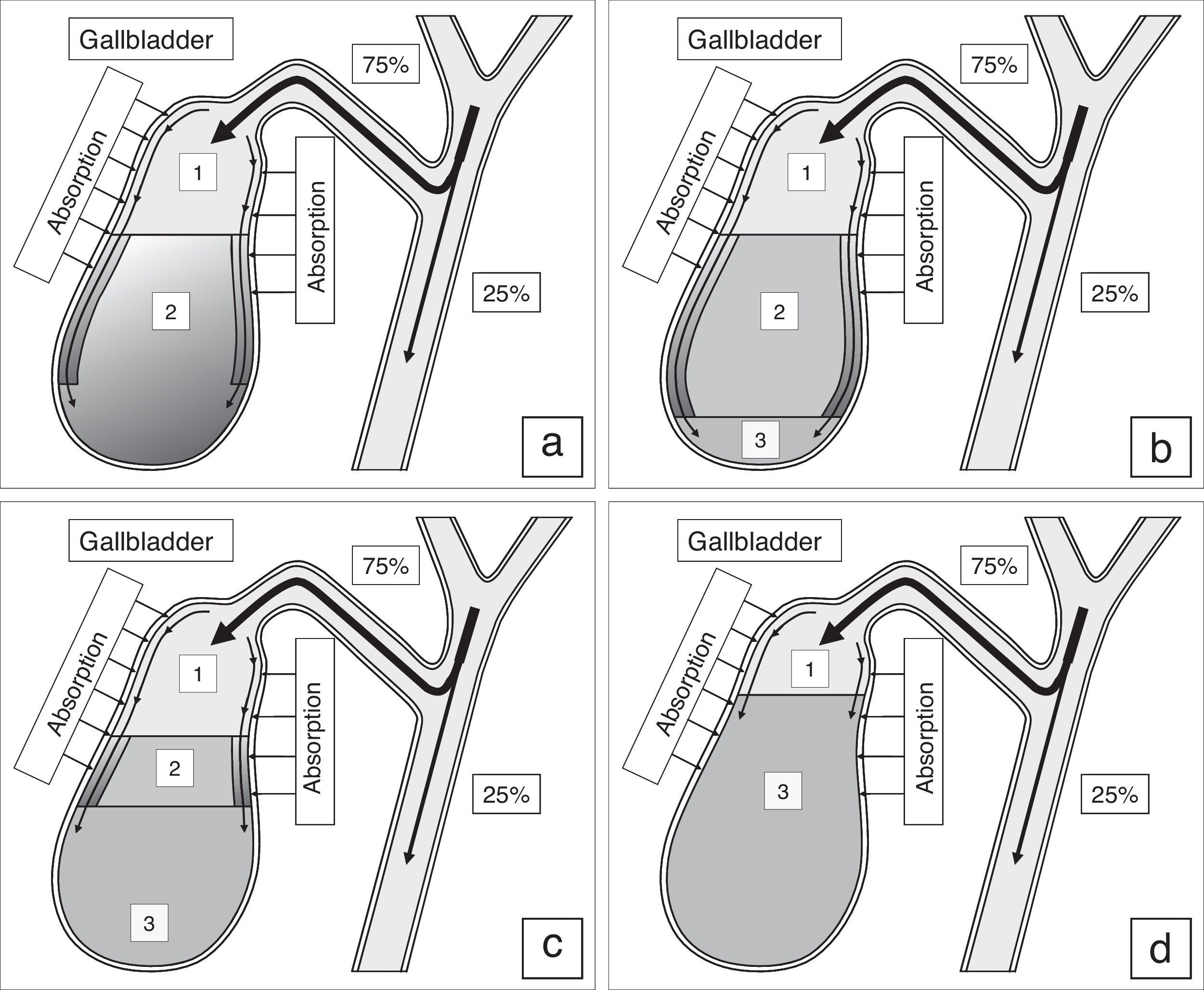
Conceptual model of gallbladder bile formationConsidering the fact that the detailed structuring of the process of hepatic bile entering the gallbladder has not been worked out, we have introduced two new terms into practice: the “active” and “passive” passages of the hepatic bile. The “active” passage depends on the ejection volume of the gallbladder after a meal or during the interdigestive period. The “passive” passage is connected with the rate of water absorption in the gallbladder. Hence the rate of the hepatic bile entering the gallbladder contains both the “active” and “passive” passages. During the “active” passage only one volume (out of 6–9) of the hepatic bile enters against 5–8 volumes during the “passive” passage. The rate of hepatic bile entering the gallbladder depends on the rate of water absorption by the gallbladder mucosa (r=+0.99, p<0.001).24 The rate of water absorption by the gallbladder mucosa is up to 100–250μl/min; sometimes it may increase up to 500μl/min.4 The rate of hepatic bile entering the gallbladder is 75% of the basal secretion of hepatic bile.24 It is indirectly confirmed by the fact that 78±10% of the bile acids from the total bile acid pool are accumulated in the gallbladder.25 The concentration of total bile acids in the gallbladder bile depends on the rate of bile acids from the hepatic bile entering the gallbladder (r=+0.87, p<0.001).24 Detailed structuring of the process of hepatic bile entering the gallbladder suggests that 83–89% of the bile acids contained in the gallbladder bile enters during the “passive” passage, and only 11–17% of the bile acids enter during the “active” passage. Hence, the “passive” passage of the hepatic bile into the gallbladder plays an important role in the mechanism of gallbladder bile formation (Fig. 1a).
 15–20min after intravenous instillation of contrast; (b) 30–40min after intravenous instillation of contrast; (c) 1.5–2.0h after intravenous instillation of contrast; (d) 2.5–3.0h after intravenous instillation of contrast. 1 – Contrasting unconcentrated hepatic bile; 2 – non-contrasting concentrated gallbladder bile; 3 – contrasting concentrated gallbladder bile.")
Process of gallbladder bile formation in healthy humans from the data of dynamic intravenous cholecystography. (a) 15–20min after intravenous instillation of contrast; (b) 30–40min after intravenous instillation of contrast; (c) 1.5–2.0h after intravenous instillation of contrast; (d) 2.5–3.0h after intravenous instillation of contrast. 1 – Contrasting unconcentrated hepatic bile; 2 – non-contrasting concentrated gallbladder bile; 3 – contrasting concentrated gallbladder bile.
Normally the filling process of the gallbladder after the intravenous introduction of X-ray contrast is characterized by some regular features.26 During the first 15–20min the gallbladder bile has two layers: the upper contrasting and the lower non-contrasting (Fig. 1a). The legible border between them is situated horizontally. During the 30–40th minute, the upper layer contrasting the bile near the wall thickens, its density grows because of the presence of heavy iodine atoms, and exceeds the density of the non-contrasting concentrated bile. In addition, the “heavy” layers of the contrasting bile begin to trickle down along the walls, as if flowing around the non-contrasting concentrated bile, and accumulate at the fundus (Fig. 1b). The gallbladder shadow becomes three-layered: the contrasting, but unconcentrated bile is above; the concentrated, but non-contrasting bile is underneath; and the contrasting and concentrated bile is beyond the lower part of gallbladder. The boundary between them is legible and it does not change if the patient moves. The quantity of the concentrated contrasting bile at the fundus of the gallbladder increases gradually, and the upper boundary of the lower layer rises (Fig. 1c). The gallbladder shadow gains homogeneity 2.5–3.0h after the contrast medium is introduced (Fig. 1d).26
Therefore, in a nighttime fasting state or in an interdigestive state, the absorption of water by the infundibulum mucosa of the gallbladder plays the leading role in gallbladder bile formation (unpublished data).
Mechanism of gallbladder bile formationTwo points in the process of gallbladder bile formation should be distinguished: (1) in a fasting stomach and (2) after postprandial gallbladder emptying.24 The absorbing and concentrating functions determine the mechanism of the gallbladder bile formation.
The rate of biliary cholesterol absorption by the gallbladder mucosa depends on the concentration of the cholesterol in the gallbladder bile (r=+0.60, p<0.001).7–9 Taking into account the fact that the mixed (bile acids-phospholipid-cholesterol) micelles are not absorbed by the gallbladder mucosa, cholesterol can be absorbed as monomers or with phospholipid vesicles.7–12,24,27–29 The solubility of anhydrous cholesterol monomers in water is 0.013nmol/ml; in the intermicellar phase it is 0.260nmol/ml, while in the phospholipid vesicles it is 5.5μmol/ml.30–37 Therefore, it will be absorbed with the phospholipid vesicles to a greater degree (99.9%), in accordance with the solubility of the anhydrous cholesterol. The phospholipid vesicles can be absorbed by the gallbladder mucosa in different ways.7–12,24,27–29 And so, the greater the absorption of vesicular cholesterol by the gallbladder mucosa is, the lower the concentration of the cholesterol in the phospholipid vesicles of the gallbladder bile.
The concentration function of the gallbladder is to accumulate the bile acids from the hepatic bile in the gallbladder; it depends on the rate that the bile acids from the hepatic bile enter the gallbladder and the rate of water absorption by the gallbladder mucosa. This function also determines the concentration of the total bile acids and the formation of mixed biliary micelles in the gallbladder bile. In the hepatic bile 40–80% of biliary cholesterol is in phospholipid vesicles and 20–60% of it is in mixed biliary micelles.35–37 The gallbladder, concentrating the bile acids, forms mixed biliary micelles and raises the level of biliary cholesterol in them up to 80–100%.35–37 Therefore, the greater the absorption of water by the gallbladder mucosa is, the greater the passage of the hepatic bile acids to the gallbladder, and the higher the concentration of total bile acids in the gallbladder bile.
Thus, the high concentration of the total bile acids and the low concentration of cholesterol in phospholipid vesicles result in the low cholesterol saturation index in the gallbladder bile (less than 1.0), which determines the stability of micellar carriers of biliary cholesterol and prevents the cholesterol monohydrate crystals from precipitating.
Fate of the absorbed vesicular biliary cholesterolPart of the absorbed cholesterol may be esterified in the epithelial cells of the gallbladder mucosa by means of ACAT. Normally the activity of ACAT makes up 92±23pmol/min per mg protein and is 8–9 times as high as that of the liver microsomes (11±2pmol/min per mg protein).38 There is a positive correlation between the concentration of the cholesterol in the gallbladder bile and ACAT in the microsomes of the gallbladder mucosa (r=+0.42, p<0.05). Cholesterol is also synthesized in the microsomes of the gallbladder mucosa, and the activity of HMG-CoA reductase in them makes up 28±6pmol/min per mg protein. But it is 4 times lower than that of the liver microsomes (120±40pmol/min per mg protein). The concentration of free cholesterol in the microsomes of the gallbladder mucosa (206±9nmol/min per mg protein) is 4 times higher than that in the liver microsomes (55±3nmol/min per mg protein), while the concentration of esterified cholesterol (34±5nmol/min per mg protein) is 3.5 times higher (9±1nmol/min per mg protein).38 Taking into account the low activity of HMG-CoA reductase and the high activity of ACAT in the microsomes of the gallbladder mucosa, as well as the positive correlation between the level of cholesterol in the gallbladder bile and the microsomes of the gallbladder mucosa (r=+0.75, p<0.01), the higher concentration of free cholesterol in the epithelial cell microsomes of the gallbladder mucosa may result only from the excessive absorption of the biliary vesicular cholesterol. It corroborates the fact that the rate of the biliary vesicular cholesterol entering the epithelial cells of the gallbladder mucosa is 4 times greater than that of the hepatocytes.
By analogy with the ileum, the removal of the absorbed vesicular cholesterol and phospholipids from the gallbladder wall may be carried out by means of HDL and/or VLDL.39–43 It was shown in vitro that HDL is able to extract the excess cholesterol from the cholesterol-saturated phospholipid vesicles and dissolve the cholesterol crystals.44,45 It is possible that the removal mechanism of the absorbed vesicular cholesterol and phospholipids by means of HDL may be prevalent and it may be determined by the concentration of serum HDL and the rate of the arterial bloodstream in the gallbladder wall.46,47 In the gallbladder wall, through its interrelation with phospholipid vesicles, HDL will extract biliary vesicular cholesterol and phospholipids and, in the bloodstream, they will first enter the gallbladder vein, then the portal vein, and from there they will enter into the liver. This supposition is indirectly corroborated by the negative correlative connection between CSI and the total cholesterol level (TCh) (r=−0.65, p<0.05) and serum Ch-HDL (r=−0.62, p<0.05) in practically healthy women.48 Small quantities of VLDL synthesize in the gallbladder mucosa.49 The number of apoproteins B, C-II and C-III absorbed by the gallbladder mucosa can make up 84–91% of the total quantity of apoproteins B, C-II and C-III, entering the gallbladder with the hepatic bile.50 In addition, taking into account that the mucosa of the gallbladder absorbs phospholipid vesicles, the apoproteins B, C-II, and C-III can interrelate with serum HDL, LDL, and VLDL in the gallbladder wall below the epithelium.
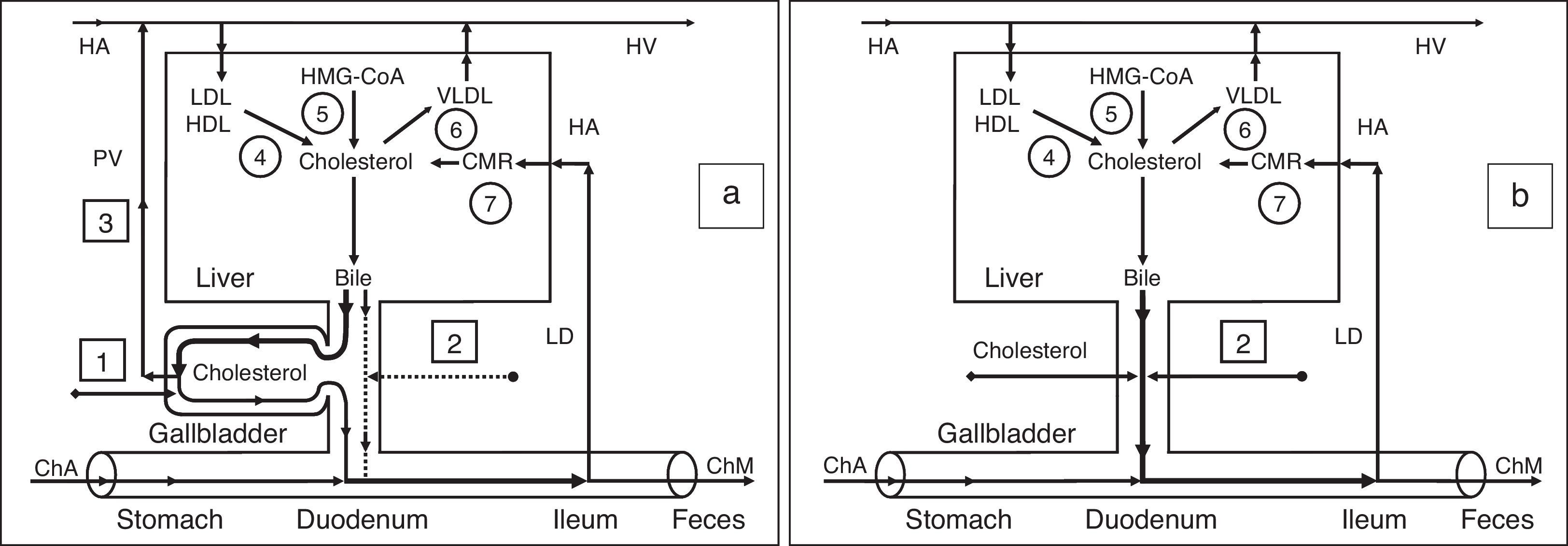
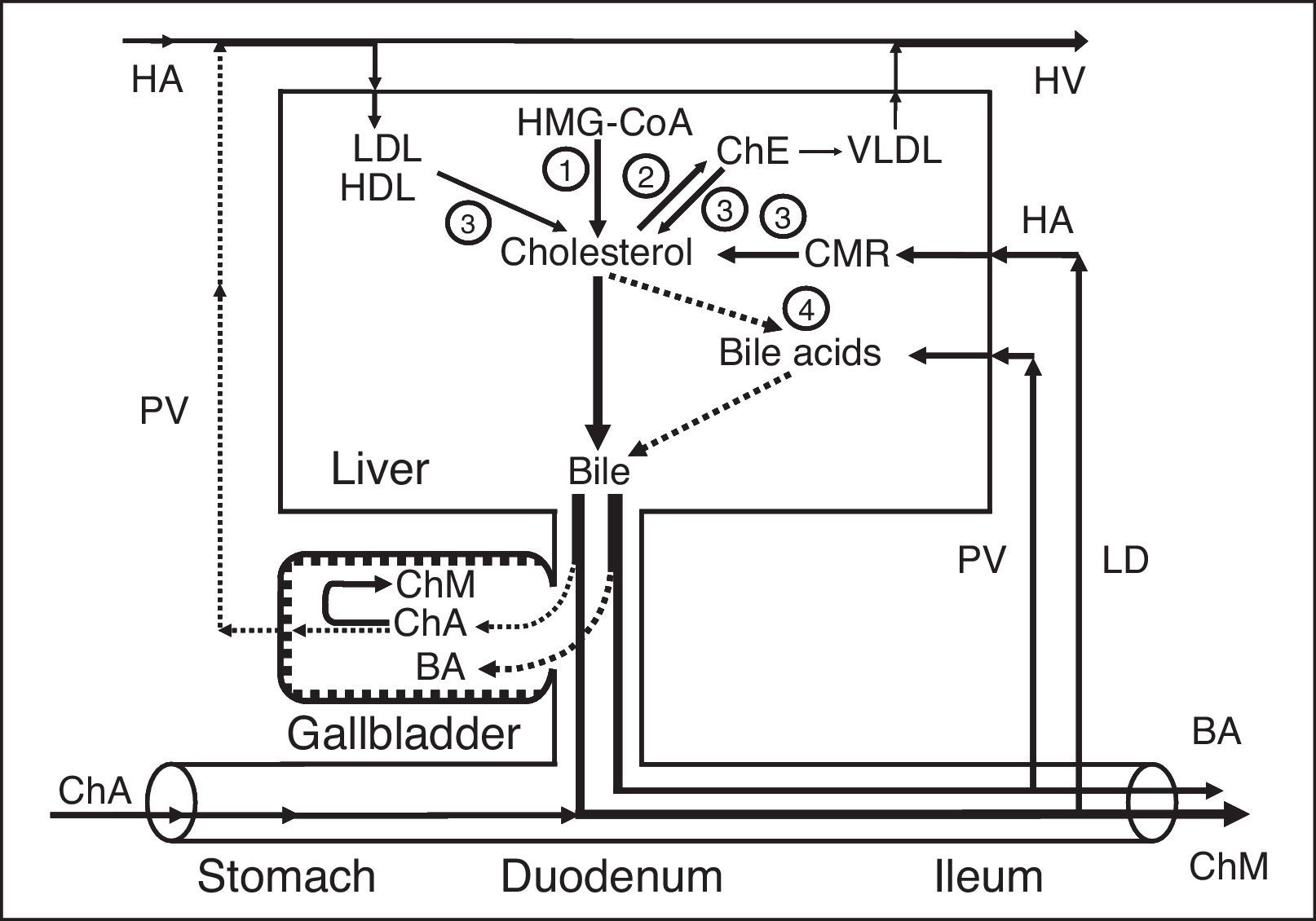
The absorbed vesicular cholesterol of the gallbladder mucosa, interrelated to blood lipoproteins, can enter the liver or the peripheral bloodstream through the portal vein (Fig. 2a).
 and patients after cholecystectomy (b). 1 – gallbladder-dependent output of biliary cholesterol; 2 – gallbladder-independent output of biliary cholesterol; 3 – gallbladder-hepatic circulation of biliary cholesterol; 4 – hydrolysis of cholesterol esters that entered the hepatocytes with HDL and LDL; 5 – synthesis of cholesterol; 6 – synthesis of cholesterol esters for VLDL; 7 – hydrolysis of cholesterol esters that entered the hepatocytes with CMR. HMG-CoA–3-hydroxy-3-methylglutaryl coenzyme A reductase; HDL, high density lipoprotein; LDL, low density lipoprotein; VLDL, very low density lipoprotein; CMR, chylomicron remnants; Ch, cholesterol; ChA, cholesterol anhydrous; ChM, cholesterol monohydrate; HA, hepatic artery; HV, hepatic vein; PV, portal vein; LD, lymphatic duct.")
Exchange of cholesterol in healthy humans (a) and patients after cholecystectomy (b). 1 – gallbladder-dependent output of biliary cholesterol; 2 – gallbladder-independent output of biliary cholesterol; 3 – gallbladder-hepatic circulation of biliary cholesterol; 4 – hydrolysis of cholesterol esters that entered the hepatocytes with HDL and LDL; 5 – synthesis of cholesterol; 6 – synthesis of cholesterol esters for VLDL; 7 – hydrolysis of cholesterol esters that entered the hepatocytes with CMR. HMG-CoA–3-hydroxy-3-methylglutaryl coenzyme A reductase; HDL, high density lipoprotein; LDL, low density lipoprotein; VLDL, very low density lipoprotein; CMR, chylomicron remnants; Ch, cholesterol; ChA, cholesterol anhydrous; ChM, cholesterol monohydrate; HA, hepatic artery; HV, hepatic vein; PV, portal vein; LD, lymphatic duct.
We have called the biliary cholesterol path (blood [lipoproteins]→liver [hepatic bile−phospholipid vesicles]→gallbladder [absorption of vesicular cholesterol]→portal vein [lipoproteins]→liver or blood) the “gallbladder-hepatic circulation of biliary cholesterol” (Fig. 2a).
The detailed structuring of these processes provides an opportunity to connect the excretory function of the liver and the absorption and evacuation functions of the gallbladder with the level of serum cholesterol.
Outflow of biliary cholesterol into the duodenumFor understanding the processes of the biliary cholesterol outflow into the duodenum, we have introduced two new terms, namely: the gallbladder-dependent and gallbladder-independent output of biliary cholesterol. The former depends on the ejection volume of the gallbladder and the concentration of biliary cholesterol in the gallbladder bile; the latter depends on the concentration of biliary cholesterol in the hepatic bile directly entering the duodenum (Fig. 2a). After cholecystectomy only gallbladder-independent output of biliary cholesterol to the duodenum is observed (Fig. 2b).
Interdependence between the absorption of biliary cholesterol in the gallbladder and that of the ileumIn the gallbladder, vesicular cholesterol absorbs effectively, but micellar cholesterol does not.7–12,24,27,28 The absorption of micelles in the ileum is 100 times more effective than that of vesicles.51 Hence, the greater the absorption of vesicular cholesterol in the gallbladder is, the higher the concentration of micellar cholesterol in the gallbladder bile (CSI<1.0), and the absorption of cholesterol in the ileum. And vice versa, the decrease of the vesicular cholesterol absorption in the gallbladder raises the vesicular cholesterol concentration in the gallbladder bile (CSI more than 1.0) and reduces the cholesterol absorption in the ileum. The bile acids/cholesterol ratio in the gallbladder bile may determine the ability of intestinal mixed micelles to solubilize dietary cholesterol. If this ratio is higher than 10–12:1 (CSI<1.0), solubilization increases, and if it is lower than 7–10:1 (CSI>1.0), solubilization decreases.
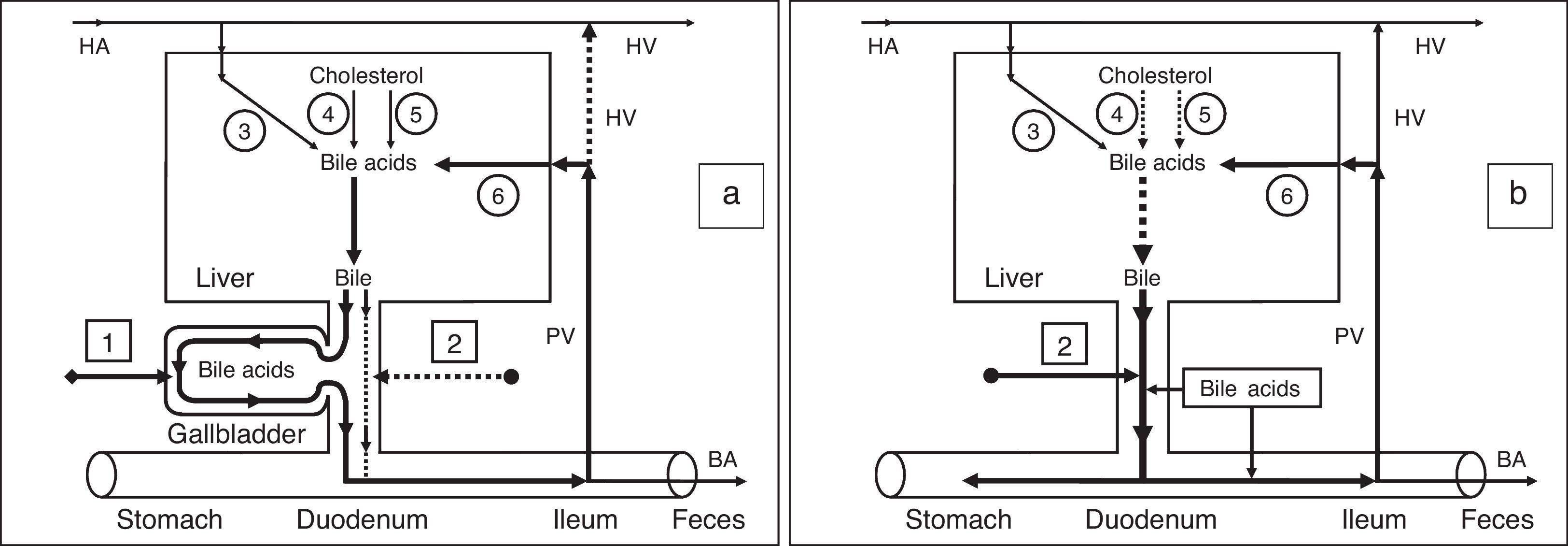
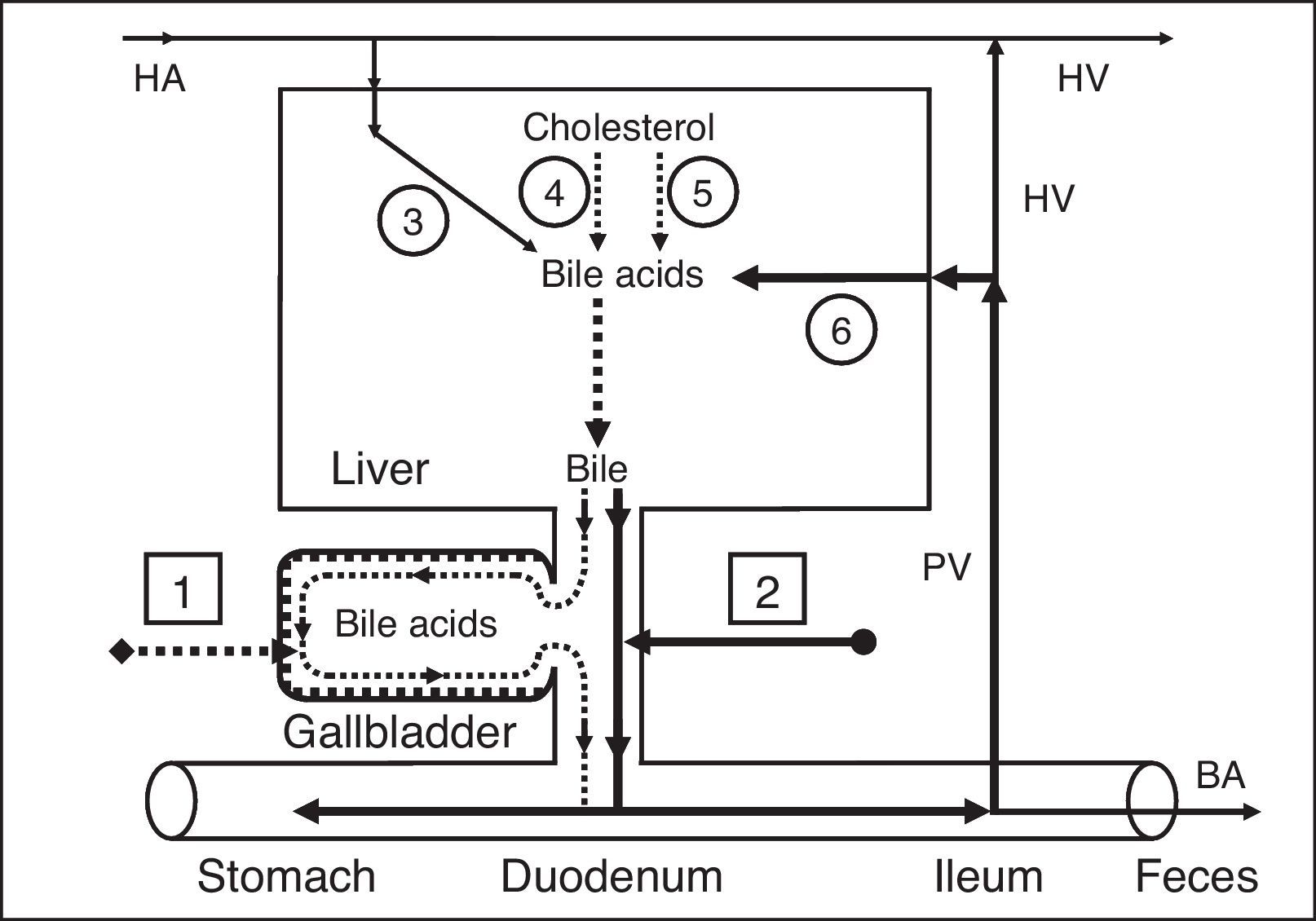
Effect of gallbladder functions on the enterohepatic circulationPart of the bile acids of the hepatic bile enters the gallbladder and is accumulated in it; the other part enters the duodenum and participates in the enterohepatic circulation. To understand these processes, we have introduced two new terms: gallbladder-dependent and gallbladder-independent enterohepatic circulation of bile acids (Fig. 3a).
 and patients after cholecystectomy (b). 1 – Gallbladder-dependent enterohepatic circulation of bile acids; 2 – gallbladder-independent enterohepatic circulation of bile acids; 3 – bile acids entering the liver through the hepatic artery; 4 – synthesis of cholic acid: cholesterol-7α-hydroxylase; 5 – synthesis of chenodeoxycholic acid: cholesterol-27-hydroxylase; 6 – bile acids entering the liver through the portal vein. BA, bile acids; HA, hepatic artery; HV, hepatic vein; PV, portal vein.")
Enterohepatic circulation of bile acids in healthy humans (a) and patients after cholecystectomy (b). 1 – Gallbladder-dependent enterohepatic circulation of bile acids; 2 – gallbladder-independent enterohepatic circulation of bile acids; 3 – bile acids entering the liver through the hepatic artery; 4 – synthesis of cholic acid: cholesterol-7α-hydroxylase; 5 – synthesis of chenodeoxycholic acid: cholesterol-27-hydroxylase; 6 – bile acids entering the liver through the portal vein. BA, bile acids; HA, hepatic artery; HV, hepatic vein; PV, portal vein.
The gallbladder-dependent enterohepatic circulation of bile acids depends on the ejection volume of the gallbladder and determines the bile acid concentration of the gallbladder bile that participates in the enterohepatic circulation. The gallbladder-independent enterohepatic circulation includes the part of the bile acids from the hepatic bile that directly enters the duodenum, but not the gallbladder. In healthy people 75–80% of bile acids participate in the gallbladder-dependent enterohepatic circulation, and only 20–25% of bile acids take part in the gallbladder-independent circulation (Fig. 3a). Therefore, the concentration function of the gallbladder is to accumulate the bile acids from the hepatic bile and exclude them from the enterohepatic circulation. The part of the bile acids participating in the gallbladder-independent enterohepatic circulation after cholecystectomy increases up to 100% (Fig. 3b). Detailed structuring of these processes enables the absorption, concentration, and evacuation functions of the gallbladder to be connected with the enterohepatic circulation of the bile acids. The rate at which water is absorbed by the gallbladder mucosa determines the passive passage of the hepatic bile from the liver into the gallbladder and the gallbladder-independent enterohepatic circulation of the bile acids.
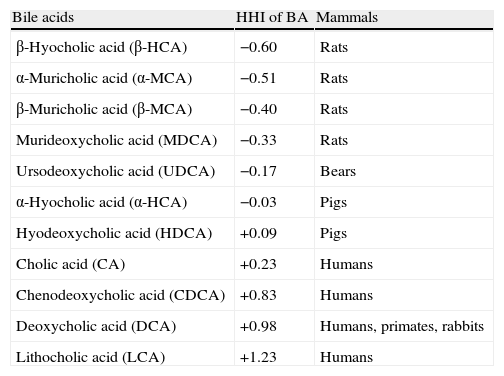
Characteristics of bile acidsAccording to the hydrophilic-hydrophobic index, the bile acids are divided into hydrophilic and hydrophobic ones (Table 1).3,52,53
Hydrophilic–hydrophobic index (HHI) of bile acids in mammals52
| Bile acids | HHI of BA | Mammals |
| β-Hyocholic acid (β-HCA) | −0.60 | Rats |
| α-Muricholic acid (α-MCA) | −0.51 | Rats |
| β-Muricholic acid (β-MCA) | −0.40 | Rats |
| Murideoxycholic acid (МDCA) | −0.33 | Rats |
| Ursodeoxycholic acid (UDCA) | −0.17 | Bears |
| α-Hyocholic acid (α-HCA) | −0.03 | Pigs |
| Hyodeoxycholic acid (HDCA) | +0.09 | Pigs |
| Cholic acid (CA) | +0.23 | Humans |
| Chenodeoxycholic acid (CDCA) | +0.83 | Humans |
| Deoxycholic acid (DCA) | +0.98 | Humans, primates, rabbits |
| Lithocholic acid (LCA) | +1.23 | Humans |
If the hydrophobic index is less than that of cholic acid (CA), the bile acids are hydrophilic; if it is more than the hydrophobic index, they are hydrophobic.3,52,53 The primary bile acids are more hydrophilic than the secondary ones, but the taurine conjugates of the bile acids are more hydrophilic than the glycine ones.3,52,53 The hydrophilic bile acids have hepatoprotective properties (muricholic [MCA]>ursodeoxycholic [UDCA]>cholic [CA]).54,55 The hydrophobic bile acids are hepatotoxic (lithocholic [LCA]>deoxycholic [DCA]>chenodeoxycholic [CDCA]>CA).3,52–57 Depending on the concentration, the hydrophobic bile acids cause cholestasis (LCA>DCA), necrosis (LCA>DCA), or apoptosis of hepatocytes (LCA>DCA>CDCA).52–57 Furthermore, DCA is cancerogenic.58 Experiments on animals showed that it causes cancer of the colon.59 The hydrophilic bile acids prevent the development of cholestasis or necrosis/apoptosis of hepatocytes (UDCA, MCA), as well as cancer of the colon (UDCA).54–57,59
In serum up to 40% of bile acids are transported with HDL and up to 15% are transported with LDL.2 The binding mechanism of bile acids with lipoproteins depends on their hydrophobic index (CDCA>DCA>UDCA>CA>7-epicholic acid).2 In the liver, there is a 60–80% bile acid uptake during one passage of portal blood.60 In earlier experiments on hamsters, it was demonstrated that the hepatic LDL uptake could influence the bile flow rate and the biliary secretion of bile acids and cholesterol.61,62 The composition and concentration of bile acids participating in the enterohepatic circulation can modulate the LDL receptor activity and the receptor-dependent LDL uptake in the liver. An increase in hydrophilic UDCA stimulates the receptor-dependent LDL uptake in the liver, but more hydrophobic CDCA decreases the LDL receptor activity.61,62 It was also shown that the addition of hydrophobic CDCA to a hypercholesterolemic diet reduces the decrease of serum HDL concentration, but the addition of hydrophilic UDCA has the opposite effect.63,64 In hepatocytes, the bile acids may inhibit the activity of HMG-CoA reductase and cholesterol-7α-hydroxylase, depending on their concentration and hydrophobic index (DCA>CDCA>CA>UDCA).52,65–67
The hydrophilic bile acids stimulate the secretion of the hepatic bile (UDCA>CA) and the hydrophobic ones lower it (LCA>DCA>CDCA).68–70 UDCA and CDCA reduce the secretion of biliary cholesterol in the hepatic bile, but CA and DCA raise it.3,68–70 In the gallbladder bile, the hydrophobic bile acids form mixed and simple micelles (DCA>CDCA>CA), but the hydrophilic bile acids form liquid crystalline lamellas (MCA>UDCA); in other words, the lower the hydrophobic index of bile acids, the lower the ability to form micelles.71–73 In the ileum, CA and CDCA raise the absorption of cholesterol, but UDCA and DCA reduce it.74–77
During the process of enterohepatic circulation in the ileum and the colon, anaerobic bacteria promote 7α-dehydroxylation of the primary bile acids (hyocholic acid [HCA], MCA, CA, CDCA) and the formation of the secondary bile acids (hyodeoxycholic acid [HDCA], murideoxycholic acid [MDCA], DCA, LCA).3,52,78,79 The secondary bile acids are more hydrophobic than the primary ones (HDCA>HCA, MDCA>MCA, DCA>CA, LCA>CDCA).3,52,53 The secondary bile acids are usually poorly absorbed in the ileum and the colon and are excreted with feces.3,52,53
The protective role of the gallbladderMammalsIn mammals (rats) that do not have a gallbladder, only the hydrophilic hepatoprotective bile acids are synthesized; as for the secondary hydrophobic hepatotoxic bile acids, they are formed in small quantities or are poorly absorbed in the ileum and the colon (Table 2).
Presence of gallbladder and type of bile acids.
| Mammals | Presence of gallbladder | Type of bile acids or bile alcohols |
| Rats | No | Hydrophilic bile acids |
| Camels | No | ? |
| Deer | No | ? |
| Elephants | No | Hydrophilic bile alcohols |
| Rhinoceros | No | Hydrophilic bile alcohols |
| Whales | No | Hydrophilic bile alcohols |
| Bears | Yes | Hydrophilic bile acids |
| Rabbits | Yes | Hydrophobic bile acids |
| Primates | Yes | Hydrophilic and hydrophobic bile acids |
| Humans | Yes | Hydrophilic and hydrophobic bile acids |
Since a long stagnation period in the gallbladder is thought to promote the formation of gallstones, the gallbladder may be absent in mammals that can manage without food and water for long periods of time (camels, deer).4 Since the size of the gallbladder must be proportional to the size of the liver, the gallbladder may be absent in big mammals (elephants, rhinoceros, whales) because of their anatomical peculiarities.4 In these mammals there is a considerable synthesis of bile alcohols, which are poorly solubilized cholesterol.2 In mammals that have a gallbladder (humans, monkeys, rabbits), both the hydrophilic and hydrophobic bile acids may be synthesized. The secondary hydrophobic hepatotoxic bile acids can be formed in large quantities, but they are poorly absorbed in the ileum and the colon.2–4 Since a long stagnation period in the gallbladder may promote the formation of gallstones, only the hydrophilic bile acids are synthesized in mammals that have long periods of hibernation (bears), but the secondary bile acids are also hydrophilic.2–4
Therefore, the basic role of the gallbladder in mammals in which hydrophobic hepatotoxic bile acids are synthesized or formed, is the protection of the liver from their effect by means of bile acid accumulation in the gallbladder and lowering the number of enterohepatic circulation cycles. The mammals in which the hydrophobic hepatotoxic bile acids are synthesized or formed must have a gallbladder. Those mammals in which the hydrophilic hepatoprotective bile acids are synthesized and the hydrophobic hepatotoxic bile acids are formed in small quantities, may manage without one. The mammals in which bile alcohols are synthesized in large amounts, do not have a gallbladder.
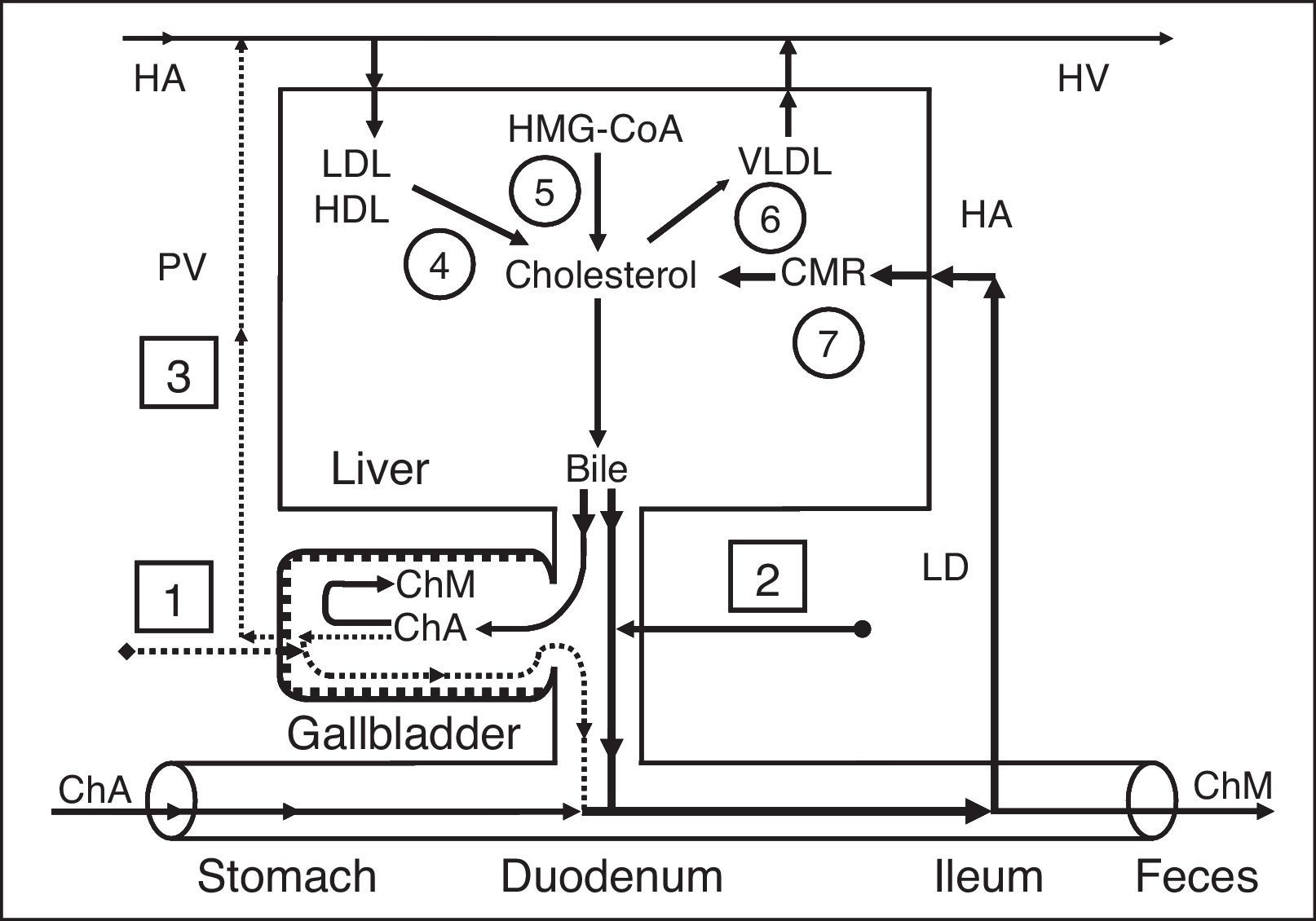
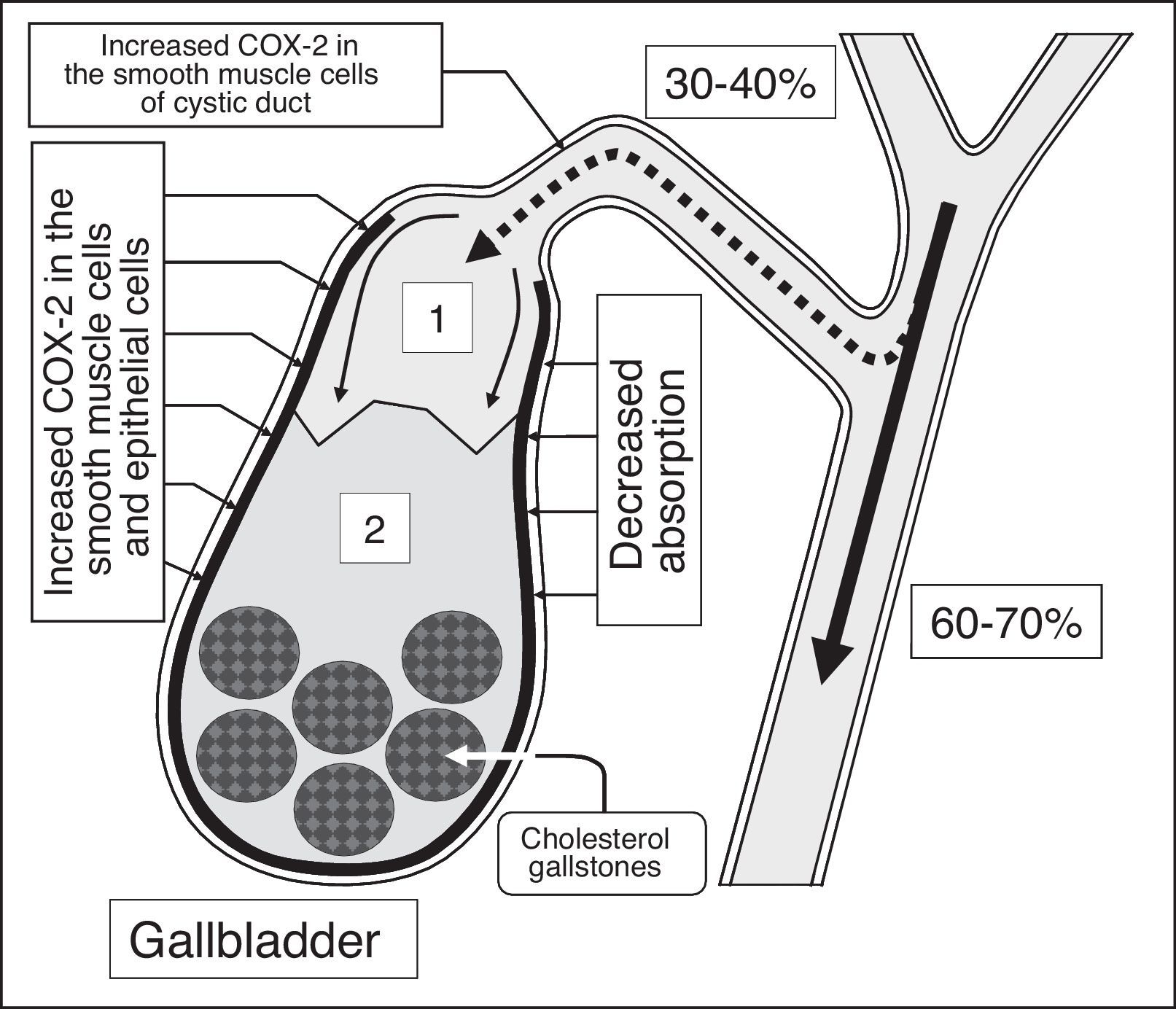
HumansIn humans, the formation of cholesterol gallstones is promoted by a decrease in absorption (a decrease in water and phospholipid vesicle absorption), concentration (a decrease in total bile acid concentration in gallbladder bile), and evacuation (a decrease in the gallbladder-dependent output of biliary cholesterol) functions and by an increase in the gallbladder's secretion function (hypersecretion of glycoprotein mucin by the gallbladder mucosa) (Fig. 4).80 A decrease in the rate of water absorption in the gallbladder wall limits the “passive” passage of the hepatic bile into the gallbladder and increases the hepatic bile passage into the duodenum (Fig. 5).6,24,80 A decrease in the evacuation function of the gallbladder reduces the “active” passage of the hepatic bile into the gallbladder.81,82 This process is accompanied by a decrease in the total bile acid concentration and an increase in the biliary cholesterol concentration in phospholipid vesicles and it also promotes an increase in the amount of time needed for the precipitation of cholesterol monohydrate crystals and the formation of cholesterol gallstones (Fig. 6).83–87 The excessive hepatic bile passage from the liver into the duodenum increases the frequency of the gallbladder-independent enterohepatic circulation of bile acids.

Cholesterol exchange in patients with chronic acalculous cholecystitis and chronic calculous cholecystitis. 1 – gallbladder-dependent output of biliary cholesterol; 2 – gallbladder-independent output of biliary cholesterol; 3 – gallbladder-hepatic circulation of biliary cholesterol; 4 – hydrolysis of cholesterol esters that entered the hepatocytes with HDL and LDL; 5 – synthesis of cholesterol; 6 – synthesis of cholesterol esters for VLDL; 7 – hydrolysis of cholesterol esters that entered the hepatocytes with CMR. ChA, cholesterol anhydrous; ChM, cholesterol monohydrate; HA, hepatic artery; HV, hepatic vein; PV, portal vein; LD, lymphatic duct.


Exchange of cholesterol and bile acids in patients with chronic acalculous cholecystitis and chronic calculous cholecystitis. 1 – synthesis of cholesterol; 2 – synthesis of cholesterol esters for VLDL; 3 – hydrolysis of cholesterol esters that entered the hepatocytes with HDL and LDL, and hydrolysis of cholesterol esters that entered the hepatocytes with CMR; 4 – synthesis of bile acids. ChE, cholesterol esters; ChA, cholesterol anhydrous; ChM, cholesterol monohydrate; BA, bile acids; HA, hepatic artery; HV, hepatic vein; PV, portal vein; LD, lymphatic duct.
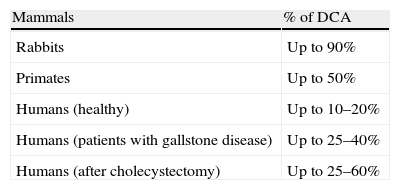
The gallbladder-independent enterohepatic circulation of bile acids in patients with cholesterol gallstone disease (CGD) or after cholecystectomy is increased (Figs. 3b and 7). It results in: (1) an increase in the hydrophobic hepatotoxic DCA formation and its accumulation in hepatocytes, (2) the formation of morphological changes in the liver (non-specific reactive hepatitis), and (3) the appearance of cholestasis (Table 3).26,88,89

Enterohepatic circulation of bile acids in patients with chronic calculous cholecystitis. 1 – Gallbladder-dependent enterohepatic circulation of bile acids; 2 – gallbladder-independent enterohepatic circulation of bile acids; 3 – bile acids entering the liver through the hepatic artery; 4 – synthesis of cholic acid: cholesterol-7α-hydroxylase; 5 – synthesis of chenodeoxycholic acid: cholesterol-27-hydroxylase; 6 – bile acids entering the liver through the portal vein. BA, bile acids; HA, hepatic artery; HV, hepatic vein; PV, portal vein.
The risk of cancer of the liver, the pancreas, the small intestine, and the colon increases as well.58,90–97 The increases in DCA, participating in the enterohepatic circulation, and other toxic agents in the hepatic bile can result in chronic pancreatitis and duodenogastric reflux.98–101
Hence, the basic role of the gallbladder in humans is a protective one. The gallbladder decreases the formation of the secondary hydrophobic hepatotoxic bile acids (DCA and LCA) by accumulating the primary bile acids (CA and CDCA) in the gallbladder and by reducing their concentration in the gallbladder-independent enterohepatic circulation, thereby protecting the liver, the mucosa of the stomach, the gallbladder, and the colon from their effect. In addition, the increase in the enterohepatic circulation cycles in humans can determine the greater enterohepatic circulation of estrogens, progesterones, and the formation of their active metabolites: 1. (a) 16α-hydroxyestrone (it activates proliferation and induces breast cancer), (b) 4-hydroxyestrone (proliferation, cancer), and (c) 2-hydroxyestrone (it stimulates fat accumulation in the human body); 2. (a) pregnenolone (inflammation, cholestasis) and (b) pregnanediol (inflammation, cholestasis).102–105
The excessive formation of these “active” metabolites and DCA most likely determines the increased risk of cancer in various sites.96,97 A total of 9.4% of the patients with gallstones and cholecystectomized patients have presented with cancer at different sites (cancer of the liver, the pancreas, the colon, and the small intestine, and breast cancer in women).96,97 For women who underwent cholecystectomy under the age of 50 (i.e. before menopause), the risk of colon cancer was higher than for those who underwent cholecystectomy above the age of 50.90 Estrogens intensify the cancer effect of hydrophobic DCA.106 The concentration of total bile acids in blood serum is 3 times higher and the risk of intrahepatic cholestasis is 2.5 times higher in cholecystectomized pregnant women (19%) than in non-cholecystectomized pregnant women.107 Children born without a gallbladder have liver function disturbances and lag behind in physical development.26 It would have been extremely difficult for humans to have evolved without a gallbladder.
The role of the gallbladder and gallbladder bile in digestionThe evacuation volume of the gallbladder depends on the quality and quantity of accepted food. The gallbladder is emptied to a greater extent when fatty food is accepted.4 Since the gallbladder contracts in 5–20min after food is available in the stomach, and “the gastric chyme” moves from the stomach into the duodenum only 1–3h later, the role of the gallbladder bile in digestion may be insignificant. The gallbladder bile, coming into the duodenum, stimulates intestinal peristalsis and promotes the cleaning of the intestine for “a new gastric chyme”.
The hepatic and gallbladder bile volumes and the bile acid concentration, participating in the first full cycle of gallbladder-dependent and gallbladder-independent enterohepatic circulation, determine the bile acid-stimulated secretion of the hepatic bile that participates to a greater extent in digestion.
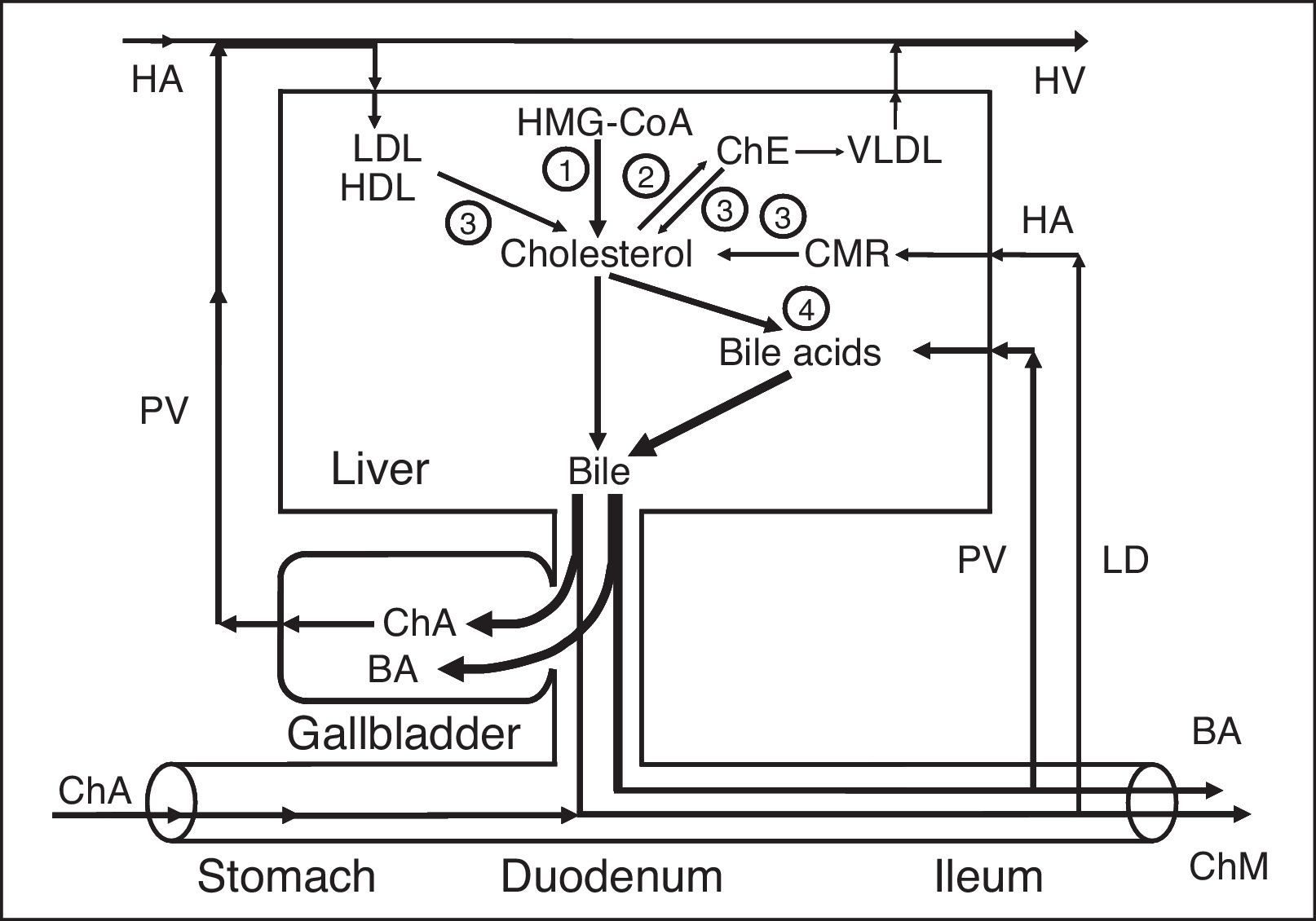
ConclusionThus, the basic role of the gallbladder in humans is to protect the liver, the mucosa of the stomach, the gallbladder, and the colon from the effect of hepatotoxic, hydrophobic bile acids, as well as to regulate the serum lipid level (Fig. 8).1–3,52–56,59,108,109

Exchange of cholesterol and bile acids in healthy humans. 1 – synthesis of cholesterol; 2 – synthesis of cholesterol esters for VLDL; 3 – hydrolysis of cholesterol esters that entered the hepatocytes with HDL and LDL, and hydrolysis of cholesterol esters that entered the hepatocytes with CMR; 4 – synthesis of bile acids. ChE, cholesterol esters; ChA, cholesterol anhydrous; ChM, cholesterol monohydrate; BA, bile acids; HA, hepatic artery; HV, hepatic vein; PV, portal vein; LD, lymphatic duct.
This gallbladder bile formation model provides a better understanding of the causes of the diseases of the hepatobiliary zone. It also allows us to visualize various trends in their treatment and prevention and to anticipate the appearance of various disorders in the hepato-biliary-pancreatic-duodenal-gastric region after cholecystectomy.
Financial supportThere was no financial support involved in the elaboration of this article.
Conflict of interestThe authors have no conflict of interest in regard to the article submitted for publication.
The authors wish to thank Ms. Leya M. Kovalyova for revising the English text and the Falk Foundation e.V. for kindly presenting scientific information.
Some parts of this work were presented at the Falk Symposium No. 84 “Bile Acids-Cholestasis-Gallstones – Advances in Basic and Clinical Bile Acid Research” (June 9–10, 1995, Berlin, Germany), the Falk Symposium No. 93 “Bile Acids in Hepatobiliary Diseases – Basic Research and Clinical Application” (October 22–24, 1996, Freiburg, Germany), the XIX Congress of the Latin American Federation of the International College of Surgeons (November 9–12, 1997, La Paz, Bolivia), the 11th World Congress of Gastroenterology (September 6–11, 1998, Vienna, Austria), the 3rd World Congress of the International Hepato-Pancreato-Biliary Association (May 24–28, 1998, Madrid, Spain) and published as abstracts.

